

Part 8 of my review of the book: “The Big Picture: On the Origins of Life, Meaning and the Universe Itself,” by Sean M. Carroll. Part 7 is found here.
Darwinian Evolution
In his next chapter “Evolution’s Bootstraps” Carroll starts by describing Richard Lenski’s experiment, which he labels as evolutionary biology. I am thinking that Carroll must have gotten the creationist message that evolution (in the goo-to-you sense) cannot be science because there is not one experimental demonstration of any process which changes microbes into molecular biologists. I say this because he states:
“Evolution is the idea that provides the bridge from abiogenesis to the grand pageant of life on Earth today. There is no question that it’s a science: evolutionary biologists formulate hypotheses, define likelihoods of different outcomes under competing hypotheses, and collect data to update our credences in those hypotheses.” (p.273, emphasis added)
In the first sentence he uses one definition for the word ‘evolution’ (the bridge from abiogenesis to the grand pageant of life) but in the following sentence it is different (mutations and selection though not explicitly stated). After the word ‘science’ what follows implies ‘evolution’ is observable in the lab, by carrying out experimental science. This is changing of the definition is called equivocation, and demonstrates very poor logic.
Very strangely Carroll does not see the point he makes himself in terms of the weakness of equivocating on the meaning of the word evolution when he admits that chemists and physicists have an advantage over evolutionary biologists because they can perform repeated experiments in their labs. The latter defines experimental science but nowhere in his statement (above) does he indicate that the evolutionary biologist carries out an experiment that “provides the bridge from abiogenesis to the grand pageant of life”.
The data collecting and formulating of hypotheses is in relation to what they believe happened in the past. At best this is historical science, a type of forensic science that tries to unravel the sequence of unseen past events. But experimental science or operational science, which is the usual definition used for science, depends on repeatable experiments to test hypotheses. This the evolutionary biologist cannot do and he admits it.
“It would be very hard to set up a laboratory experiment to see Darwinian evolution in action, just as it would be hard to create a new universe.” (p.273) (emphasis added)
Nearly correct, but not quite! It would be not “hard” but impossible. But like all evolutionists, he then equivocates at this point saying:
“But it’s not impossible. (At least for evolution: we still don’t know how to create new universes.)” (p.273)
Then he continues to introduce Lenski’s experiment on breeding tens of thousands of generations of E. coli bacteria. And, in case you missed it, I repeat, he used equivocation. He changed the meaning of the word ‘evolution’ from that bridge to one of mutations and selection in the lab.
The selection in Lenski’s experiment is not even ‘natural’ in the sense that humans are involved by providing an “extremely specific and stable environment” (p.274). The experiment produces individuals over twice the size of the original population after 60,000 generations (over about 25 years). But the mutations must have done something that makes these bacteria less fit, and less able to survive in their natural environment because Carroll also states:
“They have become very good at metabolizing glucose. While generally decaying in their ability to thrive in more diverse nutrient environments.” (p.274)
And this is the case, the mutation(s) that give them an advantage in a specialised habitat, also make them less fit as a species and hence they are evidence against the notion of a bridge to more complex well-adapted organisms as Darwin suggested. The genes to metabolize the glucose were already there, from the beginning, as part of their normal function. Those given the specialised habitat didn’t get some new complexity or function. This is natural selection at work but it only works on existing genetic information.
Carroll then adds that the E. coli had no ability to metabolize citrates, rather than glucose, but about after 31,000 generations they developed the ability. But note first,1
… Lenski seemed to have given up on ‘evolution in the lab’ and resorted to computer modelling of ‘evolution’ with a program called Avida. … Indeed, Lenski had good reason to abandon hope. He had calculated2 that all possible simple mutations must have occurred several times over but without any addition of even a simple adaptive trait.
The comments from Bob Holmes,2 in 2003, were in relation to Lenksi’s experiment at 20,000 generations, where he saw no novelty or spontaneous complexity. So what about their ability to metabolize citrates after 31,000 generations?
In a paper published in the Proceedings of the National Academy of Science, Lenski and co-workers describe how one of 12 culture lines of their bacteria has developed the capacity for metabolizing citrate as an energy source under aerobic conditions.3
This happened by the 31,500th generation. Using frozen samples of bacteria from previous generations they showed that something happened at about the 20,000th generation that paved the way for only this culture line to be able to change to citrate metabolism. They surmised, quite reasonably, that this could have been a mutation that paved the way for a further mutation that enabled citrate utilization.1
So what has happened here? Carroll clearly writes that the E. coli developed the ability to metabolize citrate, whereas “the original bacteria had no ability to use this compound”. But this is not the case, because
the citric acid, tricarboxcylic acid (TCA), or Krebs, cycle (all names for the same thing) generates and utilizes citrate in its normal oxidative metabolism of glucose and other carbohydrates.
Furthermore, E. coli is normally capable of utilizing citrate as an energy source under anaerobic conditions, with a whole suite of genes involved in its fermentation. This includes a citrate transporter gene that codes for a transporter protein embedded in the cell wall that takes citrate into the cell. This suite of genes (operon) is normally only activated under anaerobic conditions.
It is not yet clear from the published information, but a likely scenario is that mutations jammed the regulation of this operon so that the bacteria produce citrate transporter regardless of the oxidative state of the bacterium’s environment (that is, it is permanently switched on). This can be likened to having a light that switches on when the sun goes down—a sensor detects the lack of light and turns the light on. A fault in the sensor could result in the light being on all the time. That is the sort of change we are talking about.1
So in actual fact, Carroll is wrong. The ability to digest the citrate came from existing genes and it is the case that it was the result of a defect (mutation) in at least one gene that takes citrate into the cell. No ‘evolution in the lab’ was observed.
Now because he has nothing to offer as real direct observational evidence of novel structures spontaneously forming in an organism he has to write this:
“We’ve speculated that DNA came from RNA, which in turn may have self-catalyzed its own production under the right circumstances. It’s possible that the creation of the first RNA molecule involved random fluctuations at critical points along the way. Boltzmann taught us that entropy usually increases, but there is always some probability that it will occasionally move downward.” (p.275) (emphases added)
It come down to speculation and storytelling not science. Note the words he uses, which I’ve highlighted in bold text. Finally Carroll invokes a very improbably event—a preternatural miracle.
The mathematician and cosmologist Sir Fred Hoyle once stated in various ways the extreme improbability of life forming, or even getting a single functional biopolymer, such as a protein, or, an RNA molecule as he suggests here. Hoyle said,4
“Now imagine 1050 blind persons each with a scrambled Rubik cube and try to conceive of the chance of them all simultaneously arriving at the solved form. You then have the chance of arriving by random shuffling of just one of the many biopolymers on which life depends. The notion that not only the biopolymers but the operating program of a living cell could be arrived at by chance in a primordial soup here on earth is evidently nonsense of a high order. Life must plainly be a cosmic phenomenon.”
This last part indicates that Hoyle once believed that life must have come from out space, even seeded on by some alien race, knowing that it could not have occurred by random chance on Earth. Later he made the comparison about a tornado sweeping through a junkyard and assembling a Boeing 747.
So even an RNA molecule assembling itself from biomolecules in a warm pool by random shuffling is not only contrary to know physics and chemistry it is absurd to the highest order.
As a physicist Carroll should know better than to invoke the probability of a reversal in entropy as the solution, when Mr Boltzmann also taught us that the probability also considered the chance of an event NOT happening. If it is 1 chance in 1050 of it happening that also means it is 1050 -1 chance in 1050 of it not happening. And in physics any statistic smaller than 10-50 is equal to zero. It’s impossible. It just can’t happen! But Carroll holds onto hope where hope is impossible.
“The appearance of the first self-replicating RNA molecule might just have been a matter of good luck.” (p.275, emphasis added)
It really comes down to a form of faith—faith or belief in a system that has no Creator.
“From a certain perspective, Darwin’s theory is sufficiently commonsensical that it seems almost inevitable.” (p.275)
Now that’s a scientific statement if I have ever heard one! I am being sarcastic of course.
Carroll relies now solely on the story, not science. He essentially is saying it is obvious—it must have happened! Small random variations are introduced through mutations and in a stepwise process, over eons of time, change occurs in organisms. But he offers no experimental evidence to support that claim only, Lenski’s bugs, which on closer examination did not invent any novel ability.
And Carroll fails to mention or deliberately avoids the issue of all the slightly deleterious mutations that do build up in a population of any organism over time. But instead writes:
“Variations that fortuitously improve an organism’s chances of handing down its genetic heritage will be more likely to persist than those that are harmful or neutral.” (p.275)
This statement is factually wrong—and has been scientifically proven to be wrong through both experimental observation and computer modelling.5,6
The accumulation of neutral—really slightly deleterious—mutations are a serious problem for any organism over time because they are not selected for and hence not eliminated by natural selection and thus weaken the organism, making it less fit, and eventually leads to error catastrophe in the population of that organism. This is when the overwhelming burden of mutations has accumulated in the genome of the organism that they are expressed in many bad ways, which are damaging to its survival. The Second Law here is not a friend but an inexorable force causing constant decay and loss of genetic information.
It is like repairing your car over many years by replacing the engine parts to keep it running but at the same time the whole vehicle is rusting away. That rust can be identified with the slightly deleterious mutations accumulating in that analogy. It doesn’t matter how many improvements you make, the vehicle continues to rust away. And by the way, a beneficial mutation with a novel structure, arising by chance, has never been observed. But even if they do occur it does not help. The ‘rust’ never stops.
Carroll describes genetic drift where we see the change over time in an organism due to mutations and selection but he makes no mention of this very serious problem stated above. He ends the chapter with
“Can the wonders of efficiency and ingenuity we see in biological organisms really come about from random variation plus time? (Hint: yes)” (p.278)
But offers no further evidence to support such a claim. He ignore very serious objections to the theory, but believes anyway, quoting Darwin “There is grandeur in this view of life,” from On the Origin of Species. That is, take Darwin at his word, despite what we now know about genetics and the absolute impossibility of forming a single biopolymer by random chance in a warm muddy pool. The alternative cannot be contemplated.
Searching the Landscape
Now Carroll discusses natural selection, describing it as a search algorithm and gives examples of mathematical algorithms used to search out or look for some item in a very long list. In this way he is arguing that evolution is able to search for the most suited genome of an organism to survive in a particular environment.
Now remember a genome is the sequence of the particular nucleotides on the DNA strands in the genes of the organism. The human genome contains about 3 billion nucleotides, each of which can be one of 4 possible choices, symbolise by the letters A, C, G and T. Those could be arranged in 43,000,000,000 = 102,000,000,000 possible ways. So to imagine that you could find the best adapted genome by brute force flipping all the possibilities is just plain crazy. So his claim, and that of all those evolution believers, is that “[e]volution provides a strategy” (p.280) to achieve this.
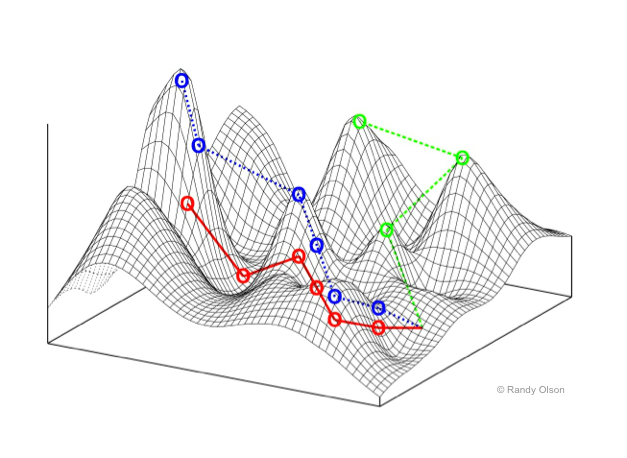
But also remember the claim is that evolution is undirected; a mindless shuffling of the code to find a more fit organism. He introduces the notion of the multidimensional fitness landscape (one dimension for each gene, for about 25,000 in humans) where evolution finds the gene most successful to survive, meaning reproduce, in the random shuffling through the landscape. See Fig. 2 (but for only 2 dimensions).
“We can think of evolution as nudging populations towards higher elevations in the fitness landscape, favouring genes that lead to more fit organisms.” (p.282)
Actually this is a faith based statement, because it is not based on actually observation. He also writes that “it can take an extremely long time for a population to climb up the landscape.” (p.282) That is like climbing up to the top of one of those hills in Fig. 2 but where there are 25,000 dimensions in the case of the human genome.
As mentioned earlier, this is contrary to known experimental evidence in terms of accumulation of mutations in an organism. The small defective mutations accumulate in the genome. They are too small for natural selection to eliminate in the next generation. If the breeding population does not get an infusion of some ‘wild’ genes from elsewhere, this leads to error catastrophe in the organism.
Because the evolutionists cannot use real empirical science is their storytelling they resort to computer modelling. Yet in those models they ignore the problem of the accumulation of all those slightly deleterious single-point mutations. In their computer models they “throw away the unsuccessful strategies and repeat the process.” (p.283), which are his words. This is not ‘mindless undirected evolution’ but ‘directed evolution’ by the programmer who has a target in mind. And really it is not even evolution; it is breeding with selection.
“If someone could show that a particular organism’s genome had high fitness within the landscape defined by its environment, but could not be ‘found’ by the strategy that evolution employs, it would decrease our credence in Darwin’s theory.” (p.287)
This leads into a discussion on Michael Behe’s concept of ‘irreducible complexity’, that an organism whose functioning involves a sequence or a set of parts that all are simultaneously required for the function, and if you took away one part it could no longer have any function. The idea that a stepwise Darwinian sequence could not be found would refute then the concept of Darwin. Behe used as an analogy a mousetrap, where if you took away one part it could not function, thus indicates design and not incremental construction by Darwin.
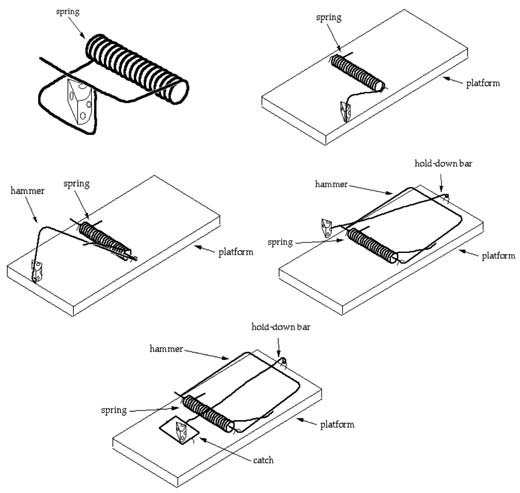
Carroll then gives a ludicrous illustration (p.288) of a sequence of images (Fig. 3) involving the progressive incremental assemblage of a mousetrap, as designed by John McDonald, which is meant to show that at each step the mousetrap is functional. It is quite laughable, in fact, if it is meant to be a true analogue of some complex biological function, like the blood-clotting cascade, which involves possibly hundreds of elements all of which need to be present for the process to work.
And Carroll does himself no credit citing Joachim Dagg, who “investigated the way actual mousetraps have changed over the years, showing that (despite being designed) they evolved gradually rather than appearing all at once.” (p.289) Surely he is joking. Mousetraps evolving? He himself says they were designed. The changes made by an intelligent mind are nothing like ‘evolution’ that Darwin imagined. Then he quotes Dagg’s words,
“All prerequisites for evolution (variation, transmission, and selection) abound in mousetrap populations.” (p.289)
Again every step of any improvement in a mousetrap was input by an intelligent mind. Variation—different design choices, transmission—different modes of passing on those designs and selection—particular choices made by an intelligent designer or user who saw the effectiveness of the mousetrap. But the primitive version was never just a single piece of wood or a single piece of metal as suggested by McDonald’s incremental evolution of a mousetrap illustration.
Next it is stated that the eye could have easily evolved. This claim has been cited many times, as if hoping that alone would make it true. And it has been in response to William Paley’s book Natural Theology, wherein he wrote that the eye implied “the necessity of an intelligent Creator”. But Carroll argues that
“Not only can eyes be explained through natural selection; they seem to have evolved separately dozens of times over history of life.” (p.289)
Did eyes evolve by Darwinian mechanisms? Author Jerry Bergman outlines the case for the eye and the impossibility of it evolving in a step-wise Darwinian fashion. Bergman writes:
The evolution of the eye has always been a dilemma for evolutionists from Darwin’s time to the present. Although Darwin, Richard Dawkins and other evolutionists have tried to explain how an eye could evolve, their solutions are clearly unsatisfactory. Many kinds of eyes exist, but no progression of eye designs from simple to complex can be produced in the natural or fossil world. Furthermore, the simplest ‘eye’, the eyespot, is not an eye but pigmented cells used for phototaxis; yet even it requires an enormously complex mechanism in order to function as a vision system.
He concludes with:
Dennett wrote that the eye lens is ‘exquisitely well-designed to do its job, and the engineering rationale for the details is unmistakable, but no designer ever articulated it.’ He concludes that its design is not real, but an illusion because evolution explains the eye without the need for a designer. This review has shown that evolution does not explain the existence of the vision system, but an intelligent designer does. The leading eye evolution researchers admit they only ‘have some understanding of how eyes might have evolved’. These explanations do not even scratch the surface of how a vision system could have arisen by evolution—let alone ‘when’.
Carroll also introduces here the part of their story that says that every time you see some structure in an ‘unrelated’ organism it evolved separately. So evolution must be common practice, that’s the implication. But it is just more storytelling, when looking at the evidence.
Like multiple evolution there is also ‘convergent evolution’!
My daughter is studying biology in university and wrote the following to me:
I just read this in my biochemistry text-book, after it was just talking about how homology (similarity in form) between different protein domains in eukaryotes means they evolved from a common ancestor.
“… in some cases however, structural similarities may be the result of convergent evolution in which a particular sequence of amino acids is independently evolved in proteins of different ancestry. E.g. the catalytic triad of proteases which occurs in a family of eukaryotic proteases with clearly homologous structures, but also in the bacterial protein, which is ancestrally unrelated to the eukaryotic enzymes but has a similar function.” (emphases added)
It is just storytelling. Whatever it suits the purpose of the story. And this is exactly the same thing that Carroll claims in his book, but this time parallel evolution because we see eyes in octopuses and human that are not evolutionary related—according to their storyline. Read Homology made simple!
The old ‘if I were God I would not have designed the human eye the way it is’ mantra is repeated.
“And the human eye, as wondrous as it is, has unambiguous flaws that would be inexcusable for a talented designer but make perfect sense in light of evolution. The nerve fibres that carry visual information to the brain are for no good reason in front of our retinas rather than behind them. … Our anatomy reflects the accidents of our evolutionary history.” (p.290) (emphasis added)
He sets himself up as some sort of judge to determine if it good or bad design. And in the case of the human eye with the optical nerve fibres in front of retina, creating a blind spot, it is apparently bad design. This is the backwardly wired retina argument. See Fig. 4. And Is our ‘inverted’ retina really ‘bad design’?
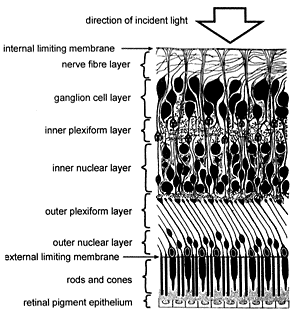
This argument was made famous by the anti-creationist atheopath Richard Dawkins in his book The Blind Watchmaker—an apparent refutation of William Paley’s book Natural Theology.
“Any engineer would naturally assume that the photocells would point towards the light, with their wires leading backwards towards the brain. He would laugh at any suggestion that the photocells might point away, from the light, with their wires departing on the side nearest the light.”
As pointed out by Jonathan Sarfati,7 Dawkins, and also hence Carroll, was actually using a theological argument rather than a scientific one.
However, ophthalmologists have denounced Dawkins’ claim repeatedly. E.g. George Marshall, the Sir Jules Thorn Lecturer in Ophthalmic Science, stated in reply to Dawkins:
‘The idea that the eye is wired backward comes from a lack of knowledge of eye function and anatomy.’
Dr Marshall explains that the nerves could not go behind the eye, because that space is reserved for the choroid, which provides the rich blood supply needed for the very metabolically active retinal pigment epithelium (RPE). This is necessary to regenerate the photoreceptors, and to absorb excess heat. So it is necessary for the nerves to go in front instead.
It’s important to note that the ‘superior’ design of Dawkins with the nerves behind the photoreceptors would require either:
- The choroid in front of the retina—but the choroid is opaque because of all the red blood cells, so this design would be as useless as an eye with a hemorrhage!
- Photoreceptors not in contact with the RPE and choroid at all—but the photoreceptors would be slow to regenerate, so it would probably take months before we could drive after we were photographed with a flashbulb, as another ophthalmologist, Joseph Calkins, points out.
- Another creationist ophthalmologist, Dr Peter Gurney in a detailed article, pointed out all the above with the RPE, but pointed out another use: extracting excess heat.
However, in the evolutionists’ fantasy world, it is only natural to take the word of evolutionists totally lacking in ophthalmology qualifications over creationist experts in ophthalmology, when the issue is ophthalmology.7
Jonathan Sarfati also commented:
Ophthalmologically ignorant evolutionists like Richard Dawkins and Kenneth Miller claim that the backward wiring of the vertebrate retina is ‘bad design’. But:
“[T]he retina of the eye has been optimised so that the sizes and densities of glial cells match the colours to which the eye is sensitive (which is in itself an optimisation process suited to our needs). This optimisation is such that colour vision during the day is enhanced, while night-time vision suffers very little.”
Evolutionary arguments about the eye are, again, solely motivated by a belief in evolution as the true history of this planet. The evolutionist has no reason to assume that there exists any rhyme or reason or purpose for the alleged changes. The biblical creationist sees creation as the product of an intelligence, the one and only Creator God, who we know as the Lord Jesus Christ (John 1:2 and Hebrews 11:3).
Click here for Part 9 of this review.
References
- Batten, D., Bacteria ‘evolving in the lab’? creation.com/bacteria-evolving-in-the-lab-lenski-citrate-digesting-e-coli
- Holmes, Bob, Bacteria make major evolutionary shift in the lab, NewScientist.com news service, 9 June 2008.
- Blount, Z.D., Borland, C.Z. and Lenski, R.E., Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli, PNAS 105: 7899–7906; published online on June 4, 2008, 10.1073/pnas.0803151105. This is Lenski’s inaugural paper as a newly inducted member of the National Academy of Sciences, USA—yet another dyed-in-the-wool atheistic evolutionist in that august body (see: National Academy of Science is godless to the core Nature survey).
- Hoyle, Fred, The Big Bang in Astronomy, New Scientist 92:521–527, 1981.
- Sanford, J., Baumgardner, J., Brewer, W., Gibson, P., and ReMine, W., Mendel’s Accountant: A biologically realistic forward-time population genetics program, SCPE 8(2):147–165, 2007. Springer-Verlag, Berlin, Heidelberg, pp. 386–392. (PDF, the program is available for download here.)
- Batten, D., Genetic algorithms—do they show that evolution works? creation.com/genetic-algorithms-do-they-show-that-evolution-works
- Sarfati, J.D., Fibre optics in eye demolish atheistic ‘bad design’ argument, Creation 31(1):45–47, December 2008; creation.com/fibre-optics-in-eye-demolish-atheistic-bad-design-argument
Follow me
- Telegram.org: @GideonHartnett
- Facebook: Gideon Hartnett
- X (Twitter): @gideon195203

To be notified by email put your email address in the box at the bottom of your screen. You’ll get an email each time we publish a new article.

Click this image to make a secure Donation (Stripe) !





